Joint Research Group Macromolecular Crystallography
Structure of the month
PNAS 2012 109(14):5235-40.
Redox-dependent Complex Formation by an ATP-dependent Activator of the Corrinoid/Iron-Sulfur Protein
Sandra Elisabeth Hennig1#, Jae-Hun Jeoung1#, Sebastian Goetzl1, Holger Dobbek1
1 Institut für Biologie, Strukturbiologie/Biochemie, Humboldt-Universität zu Berlin, 10115 Berlin, Germany
# These authors contributed equally to this work
Corresponding author: Holger Dobbek, Institut für Biologie, Strukturbiologie/Biochemie, Humboldt-Universität zu Berlin, 10115 Berlin
E-mail: holger.dobbek@biologie.hu-berlin.de
Phone: ++49-30-2093-6369; Fax: ++49-30-2093-6447
Abstract
Movement, cell division, protein biosynthesis and electron transfer against an electrochemical gradient. These and many more processes depend on energy conversions coupled to the hydrolysis of ATP. The reduction of metal sites with low reduction potentials (E0’ < –500 mV) is possible by connecting an energetical uphill electron transfer with the hydrolysis of ATP. The corrinoid-iron/sulfur protein (CoFeSP) operates within the reductive acetyl-CoA pathway by transfering a methyl-group from methyltetrahydrofolate bound to a methyltransferase to the [Ni-Ni-Fe4S4] cluster of acetyl-CoA synthase. Methylation of CoFeSP only occurs in the low-potential Co(I) state, which can be sporadically oxidized to the inactive Co(II) state, making its reductive reactivation necessary. Here we show that an open-reading frame proximal to the structural genes of CoFeSP encodes an ATP-dependent reductive activator of CoFeSP. Our biochemical and structural analysis uncovers a novel type of reductive activator, unrelated to the electron transferring ATPases found to reduce the MoFe-nitrogenase and 2-hydroxyacyl-CoA dehydratases. The CoFeSP activator is structurally homologous to actin, hexokinases and heat-shock proteins and derives its electrons from a ferredoxin-like [2Fe–2S] cluster. Complex formation between CoFeSP and its activator depends on the oxidation state of CoFeSP, which provides evidence for a novel strategy to achieve unidirectional electron transfer between two redox proteins.
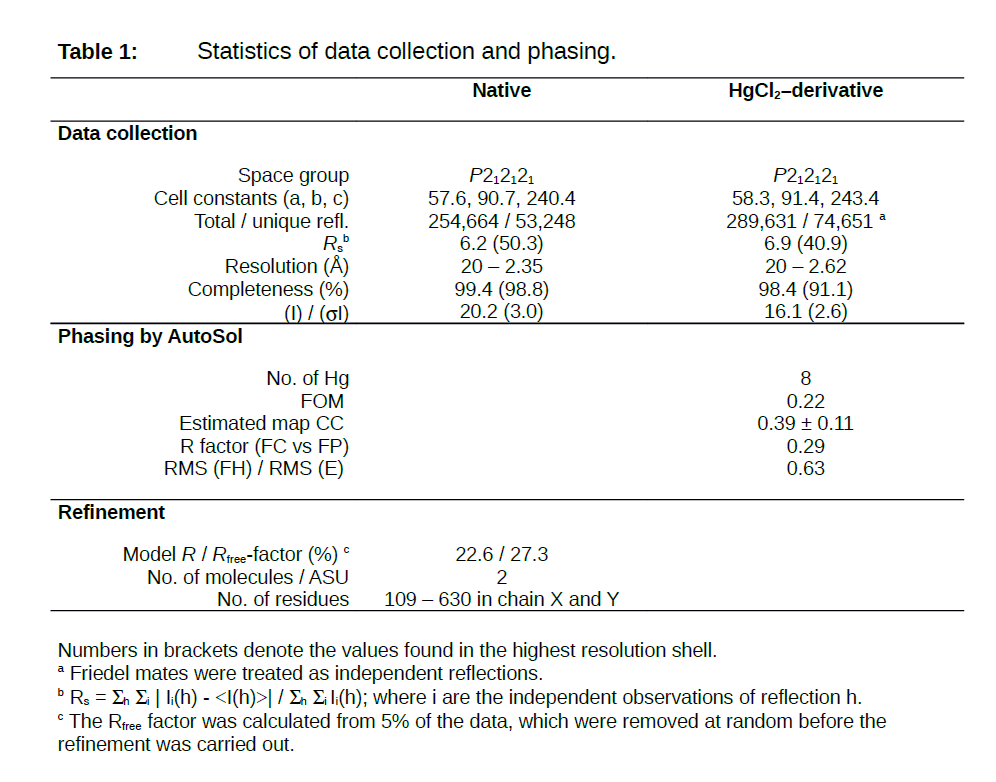
Table 1.

Figure 1.
Crystal structure of the activator. The two molecules of RACo in the asymmetric unit of the crystal are presented as cartoons. The left molecule (subunit A) contains all domains of the activator including the N-terminal domain (cyan), the linker domain (purple), the middle domain (blue) and the ASKHA domain (red). The second monomer (subunit B) is colored in grey and does not include the N-terminal domain in its structure. Magnesium and phosphate ions are indicated as green and gray spheres, respectively. The [2Fe-2S] cluster is shown as spheres colored red for iron and yellow for sulfur.
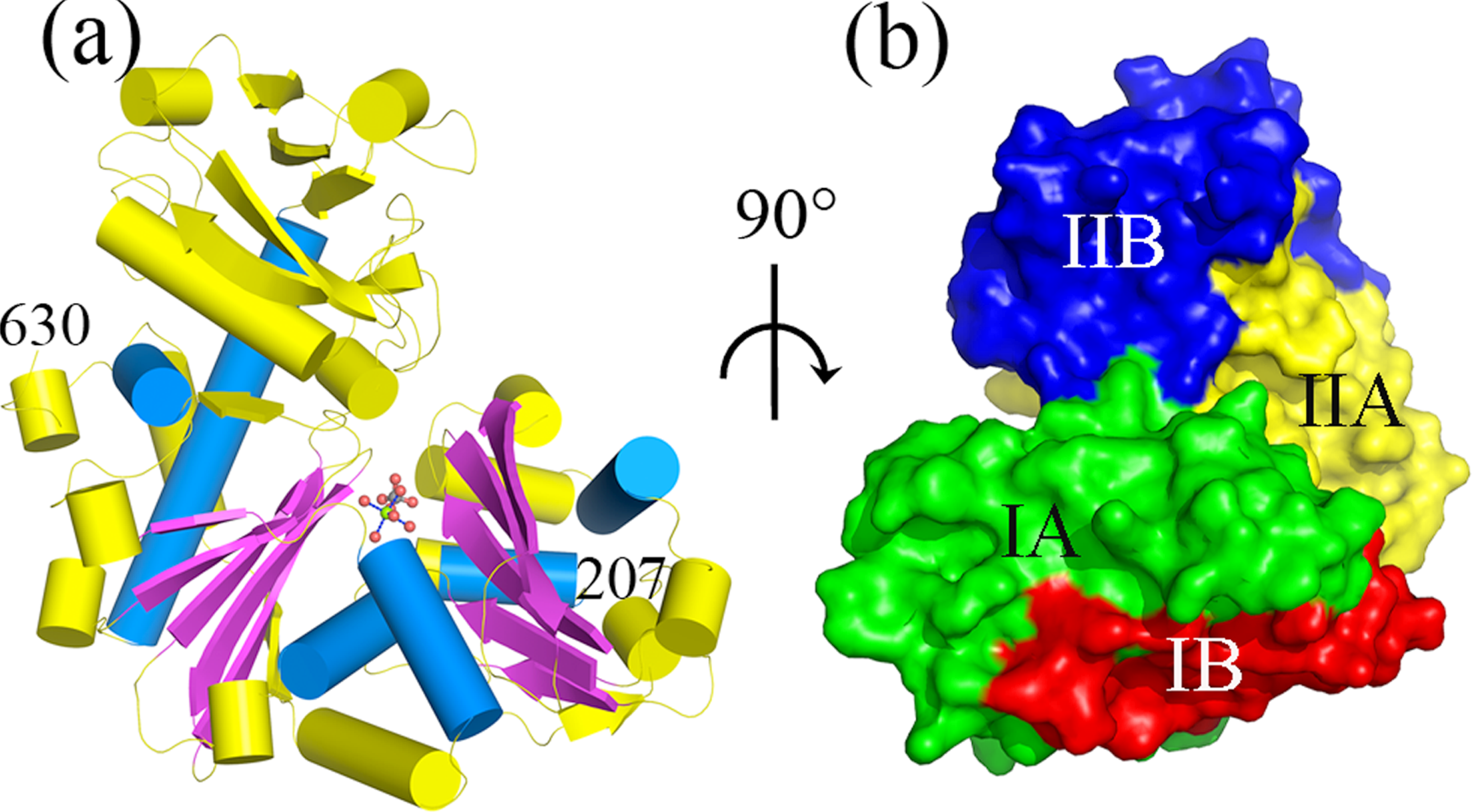
Figure 2.
Cartoon-representation of the ASKHA domain. The ASKHA domain of RACo (residues 207 630) is shown in (a). The conserved βββαβαβα topology of the ASKHA family is illustrated with βstrands in magenta and αhelices in marine. Surface representation of the four lobes of the ASKHA domain (IA, IB, IIA and IIB) in analogy to the ASKHA family proteins is given in (b).